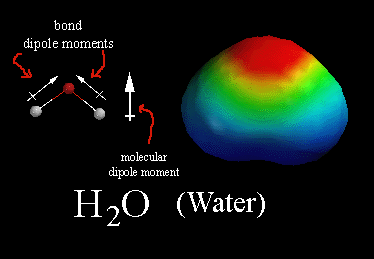elkészültek a képek (: mindent MOE-al csináltam, direkt az 1LBT.pdb-bõl, hogy bemutassam a lipáz mûködésének modelljét.
1. A megnyitott pdb fájl a két enzimmel, amint említettem:
2. A jobb oldali enzim kitörlése után, és a T80-as beszínezett szubsztrát az aktív centrumban.
3. Az fehérje megjelenítését átváltottam; az aktív centrum közelebbrõl, H-ek hozzáadása után (ezt nem tartalmazza a pdb, mert a felbontása túl kicsi volt a szerkezetfeloldásnál), valamint a két fontos szerpet játszó oldallánc, a 105-ös szerint (SER_105) és a 224-es hisztidint (HIS_224) megjelöltem
4. A fölösleges H-eket elrejtettem a láthatóság kedvéért, bejelöltem 2 távolságot
5. A katalízis elsõ lépéseinek eredménye, a Ser nukleofilizálása a His által, majd a nukleofil támadás a szubsztrátra
6. Optimalizáltam a Ser körüli részt, fõképp a kötéstávolságok miatt, bejelöltem a következõ lépést.
7. Az alkohol termék létrejötte
Az ezután következõ lépés az OH kidiffundálása és egy H2O molekula elhidrolizálja a szubsztrát enzim komplexet és kialakulnak a termékek, valamint az alapállapotú enzim. Szerintem ezen a példán is nagyon jól látszik az enzimek szerepe, hogy csökkentsék az aktiválási gátat, az által, hogy "beágyaznak" a közti állapotoknak, így kedvezõbbek lesznek a lépések.
remélem tetszett (:
azt hiszem nem itt beszéltünk az öregedésrõl, mostanában sokan kérdezik h h s mint is van ez :D
szóval én úgy látom, hogy az öregedés, az messzemenõen nem egy faktortól függ és nem is egy helyen történõ változás miatt következik be, hanem komplex események együttes eredõje.
Szerintem az ember, valamilyen oknál fogva, sokszor fel sem méri a szervezetben lejátszódó folyamatok minõségét és mennyiségét. Ilyenekre gondolok: Depurináció, ami azt jelenti, hogy a DNS nukleotidokról a bázis spontán lehidrolizál. Ez a folyamat egyetlen sejtben, napi 5000-szer megtörténhet [1], és legtöbbször kijavítódik enzimek által. Sztem az ember nincs hozzászokva ahhoz a gondolathoz, hogy a molekulák mennyire instabilak (erre utaltam fönt), és ez csak egyetlen jelenség, ott vannak a DNS szabad gyökök általi támadása (fõleg ROS, Reactive Oxygen Species).
Egy proteaszómás cikknek az olvasásakor döbbentem rá arra, hogy a fehérjéket sem értjük meg ilyen szempontból. Egy fehérje átlagos élettartama durván 1 nap, legöregebb fehérjéink pár hetesek és sok fehérje csak percekig létezik [2]. Ez megint valamiért nem annyira intuitív. A balansz fenntartása a szintézis és degradáció között létfontosságú.
Egy másik központi átalakításokat végzõ folyamat a DNS-metilezése, amelynek hatására a sejt bizonyos génjei elhalkulnak (szupresszálódnak) és így alakulhat ki a specializálódott sejt. A metilezés ezentúl, a sejt életében is, dinamikusan folyik az igények szerint. Viszont ez a folyamat sok hibalehetõséggel jár, mert a metilezésnek csak bizonyos gének ELÕTTI régiókban van értelme (ún. CpG-szigetek), ha a gének metilálódnak, az mutációkhoz vezethet és mivel a folyamat sokszor lejátszódik és a fehérjék messze nem tökéletesen végzik a munkájukat, ez gyakran meg is történik.
Még mindig nagyon kevés témát említettünk és már kezd hosszú lenni a hsz.-em :D Ha megpróbáljuk azt is hozzágondolni, hogy minden ilyen irányító folyamatok milyen sok tényezõsek, amelyek itt-ott, egyszer itt, egyszer ott sokszor kibillenek az egyensúlyból, az öregedés szükségszerûnek látszik. Én úgy látom, hogy mintha maga az élet okozna zavarást a rendszerben, amire a rendszer úgy reagál, hogy a zavarást csökkentse (Le Chatelier-elv), és meg is teszi, a zavarás megszûnik, szinuszosan idõben lecsengve.
Oh, hülyeség is, amire gondoltam, az oxigén a kettõs kötésével stabilan el van, nem kell neki egy hidrogén.
Az öregedésrõl egy ism. terj.-ben azt mondták, h a DNS javítás során keletkezõ hibák okozzák. Mivel nem volt világos számomra, a korábban írt javaslataid is ezt célozzák?
hú, bocsi h összezavartalak. a képen valóban 1 fehérjét mutat meg még a Jmol-ban is :S (ezt csak most láttam) én csak a letöltött a pdb fájllal dolgoztam és abban két fehérje van egymás mellett, gondoltam a jmol is ezt nyitja meg, de ezek szerint nem.
a hisztidin gyûrûben lévõ N-je nyúlja le a szerin protonját (: majd csinálok képeket is, amúgyis szeretnék :) akkor majd a számolásról is csinálok, amúgy a mi szubsztrátunk egy phenothiazine származék volt.
Izé, nem program az, csak az van a 1LBT linken jobb oldalt a kép fölé írva, de most meg sztem az a két kép ugyanerrõl készült, csak különbözõ szögekbõl. "itt eleve 2 fehérje van a fájlba, valamiért így jött össze" Vagy ezzel a T80-ra és a NAG-ra gondoltál?
Asszem most esett le a szubsztrát bekapcsolódása. A hisztidin észteres része lenyúlja a szerin terminális protonját, ettõl a szerin meg nukleofil lesz és a karbonilt megszereti, sp3 hibridizációt kényszerít ki és az észterre jobban áttevõdnek a kötõ elektronok. így a karbonil egy királis centrum lesz. A 3. bekezdésedben pont fordítva soroltad fel az ok-okozati viszonyokat, ez kicsit megzavart (azt hittem már értenem kéne, miért is történik, ami történik az elsõ mondatokban). Mi volt az általatok modellezett királis szubsztrát?
Úgy rémlik a Veszprémi TK-ban is van érdekesség a víz szerkezetének kutatásáról. Az jó, üdítõ volt, már épp kezdtem azt hinni, h nincsenek ott nagyobb bonyodalmak.
azt a programot nem ismerem, én a "hyperchem"-et, vagy a "Spartan"-t, vagy a "molecular operating environment"-et használtam. Egyik sem ingyenes, de neten "fellelhetõ".
T80 és NAG igen, de valszeg T80 nélkül is tudták volna kristályosítani, de ily módon megkapták az enzimet a szubsztrátjával az aktív centrumban, ami több, mint csupán az enzim.
Ha lesz idõd/kedved/stb. sztem olvasd végig, leírtam kb mindent ami érdekes az enzimrõl.
A diffrakciós képet nem szokták korrigálni a torzulások miatt, tudomásom szerint ...; legtöbb célra így is megfelelnek a modellek.
Ha a lipázt nézted (1LBT), itt eleve 2 fehérje van a fájlba, valamiért így jött össze. Ez nem egy dimer fehérje, hiszen látszik, hogy két különálló polipeptid. Itt benne van az aktív centrumba a saját szubsztrátja, amivel izolálták (T80), a fehérje adatlapján alul ezt írja is (+ az NAG molekulák amelyek a folyamat során kellhettek a fehérje mellé).
Szóval ha megkeresed az egyik polipeptidben a T80-at, akkor ezzel az aktív centrumot is megtalálod. A T80-nak van egy észter csoportja, ezt hidrolizálja el a lipáz. Ha az észter csoportot megtalálod, a közelben lesz a fehérje egyik szerin oldallánca is, amivel a nukleofil támadást végzi a szubsztrát karbonil szenére, ahogy az a szerin-hidrolázokhoz illik (: A közelben egy hisztidin oldallánc is befigyel :D Az nukleofilizálja a szerint, oly módon, hogy a terminális -OH csoportjáról leveszi a protont, és az így létrejövõ deprotonált hidroxilcsoport erõs nukleofil tulajdonságokkal rendelkezik.
Amikor a deprotonált szerint oxigénje betámad a szubsztrát karbonil szenére, az sp2 hibridállapotból átmegy sp3-ba, ez a tetrahedrális intermedier (THI), úgy hogy a karbonil kettõskötés pi elektronjai a karbonil oxigénre mennek, ezért az intermedier negatívan töltött. Az érdekesség kedvéért még hozzáteszem, hogy a fehérje úgy van kialakítva, hogy ezt a negatív oxigént poláris oldalláncok stabilizálják, ezt hívják oxianion lyuknak (oxyanion hole).
A T80 nem királis (amúgy a belõle létrejövõ THI igen). Mi Hyperchemmel kitöröltük a T80-at és helyette bemodelleztük a saját királis szubsztrátunkat, amelynek volt R és S változata, valamint a létrejövõ THI megint lehetett R vagy S, aszerint, hogy a Ser merrõl támadta. Így összesen 4 modell volt, amellyel számolni kellett. Ezt én az otthoni gépen csináltam (a számolást is), viszont csak molekulamechanikát, kvantum számításokat nem. Kb 1 nap 1 számítás, de én választom ki, hogy meddig próbálgassa (random pozícióknak számolja az energiáit).
Ezek nem olyan komoly számítások, mert kicsi a rendszer (szubsztrát+annak kb. 10 angströmös környezete), de pl. nemrég olvastam egy cikket, hogy a víz fagyását elindító nukleációt modellezték szuperszámítógépeken és úgy is hónapokig eltartott nagyon rövid idõ modellezése.
Ha netán elakadnál írj nyugodtan, leírom pontosan az aminosav számokat, hogy miket kell nézni, képeket is csinálhatok arról amiket én csináltam...
Huh és még azokkal a lehetséges torzulásokkal is számolnak legalább becslés szintjén?
A linket köszi. Jó cucc ez a JMol. Gondoltam megnézem a királis változatokat is (tényleg, ennél hány kiralitási centrum van?), de nem találtam, hogy hol lehetne.
Egyébként a számolásokhoz a spektroszkópia labor gépeit használtátok? Mert nem tom, mennyire kell ehhez számítási igény és mindenesetre ott mondták Kállay mesterék, h az õ klaszterüket szokták a kvantum kémiai számításokra használni, de mondjuk ez nem zárja ki, h más klaszter is volna, ahol számolgatnak.
de valószínûleg fellép :), ráadásul ami ennél nagyobb probléma, hogy fiziológiás körülmények között vizes közegben van és a vízmolekulák lazíthatják/feszíthetik akár az egész polipeptidet, akár annak egy kis részét, és ugyebár ez a kristályosított fehérjében nem látszik, mert csak nagyon kevés víz marad.
amúgy ha érdekel, simán felnézhetsz a www.pdb.org/-ra itt nagyon sok fehérje van és bele tudsz nézni bármibe (segítek ha kell), mi is innen szedtük a fehérjét amivel dolgoztunk, ha érdekel keress rá arra, hogy 1LBT, ez a PDB_ID-je. Az oldalon megtudod nyitni JMol-al (java-s progi), számolni azzal nem lehet, mert ahhoz gyenge. Számoláshoz kell legalább egy Hyperchem. A TDK-s dolgot meg igen, jól értetted (:
tudnak ám nem tudtam, h ilyen jó felbontást tudnak ilyen összetett molekulákkal is (pedig gondolhattam volna) ilyenkor a rács léte miatt, a másodlagos kh.-ból nem lép fel nagyon kis mértékû torzulás a szerkezetben? Bár talán azt is figyelembe lehetne venni utólag...
a TDK-sat meg jól értem, h akkor a modellszámítással különbözõ kiralitású eseteket vizsgáltatok és azon esetek közül, ahol az átmeneti molekula energiája kisebb volt, azzal a valószínûbb, h elreagál?
amúgy 2-3 angström felbontású szokott lenni egy diffrakciód kép; én TDK-n csináltam olyat a szerves tanszéken, hogy egy sztereoszelektív lipáz-t modelleztünk, és arra próbáltunk rájönni, hogy a reakció során melyik királis molekulával reagál el. Erre onnan következtettünk, hogy mekkora az energiája az enzimben kialakult átmeneti állapotú molekulának (az ún. tetrahedrális intermedier) az energiája a különbözõ esetekben.
nem számolásból csinálják a fehérjék képeit, hanem röntgen diffrakcióval :) (X-ray crystallography), ennél azt szokták mondani h a fehérjék kristályosítani a legnehezebb, mert olyan, mintha "bowling golyókat kéne ragasztószalaggal összeragasztani"; ezután a diffrakció rekonstruálása is sok idõt vehet igénybe ...
Kvantumszámításokat fehérjén végzett számolásoknál szoktak alkalmazni. Ilyenkor már megvan a számítógépes modell diffrakcióból, és pl. belemodellezel egy szubsztrátot és akkor annak az energiáját kitudod számolni akár QM modellel, akár mechanikai modellel (molekula dinamika), vagy van a kettõ között átmenet is.
Ezeknek a nagy fehérjéknek a modellezése mennyire pontos? A Schrödinger 1enlet numerikus közelítésével hosszú idõ alatt kapják meg az alakjukat, esetleg még élnek más közelítésekkel is vagy már meg sem próbálják megoldani, csak reakciók alapján és egyéb ismeretek alapján figurázzák ki a konstrukciót? Merthogy én úgy hallottam, h a nagy molekulákra való számolgatással bajok vannak és ezért is várják már a kvantumszámítógépeket.
"Én úgy vagyok, hogy nem tudom elfogadni azt a tényt, hogy öregedünk" Én ugyanígy voltam a supermanséggel. Ezért alábbhagytam az elvárásaim és már csupán az örök fiatalság hiányától szenvedek. De 1iket sem látom, h a közeljövõben elérjem, ezért nagy ám a világfájdalmam. xD
Gondolom arról hallottál már, h fonalférgeknél pl. egy gén kiiktatásával 6szor tovább éltek egészségesen.
Egy kis összefoglaló annak aki nem akarja elolvasni az egész cikket: a preoteaszóma egy olyan óriás fehérje komplex (≈2000 kDa), amely az eukarióták citoplazmájában helyezkedik el és más fehérjéket képes lebontani. A komplex által lebontandó fehérjék poli-ubikvitinnel vannak megjelölve, amely egy fehérje (ubikvitin) polimer, maga a jelölés folyamatát ubikvilációnak hívják.
Az ubikviláció során a a lebontandó fehérjék egyik lizin aminosav oldalláncának ε-aminocsoportjáhóz segédfehérjéken keresztül hozzákapcsolódik az ubikvitin karboxil terminusa, és így létrejön egy "izopeptid kötés". Ezután az ubikvitinhez további ubikvitin fehérjék kapcsolódhatnak, így hozva létre a a poliubikvitin-t a lebontandó fehérjén.
Az ubikvitin és annak funkciójának felfedezéséért 2004-ben adtak Nobel-díjat, 3 tudós között megosztva: Aaron Ciechanover, Avram Hershko és Irwin Rose-nak, akik közül Hreshko magyar származású, Izraelbe kitelepült biokémikus.
sztem tök jó, hogy beraktad (: én legalábbis örültem neki, még akkor is ha nem feltétlenül tetszett a stílusa; de miért kéne tetsszen és miért kéne nekem? (: (költõi kérdések)
engem az állatok és gépek ötvözése bizonyos szinten érdekel. Én úgy vagyok, hogy nem tudom elfogadni azt a tényt, hogy öregedünk (: Idegesít az a gondolat, hogy csak egy meghatározott, vagy egy meg nem határozott, de véges idõintervallum áll rendelkezésünkre, hogy megértsünk "dolgokat";
Nincsenek túlzottan speciális ismereteim a témáról, mert még az alapokat próbálom megérteni, de én úgy látom, hogy lényegében kétféleképpen lehetne megállítani/manipulálni az öregedést (és mind2 nagyon futurisztikus): -vagy sejtszinten kéne módosítani a rendszert, hogy stacioner állapot tudjon kialakulni (telomer regeneráció, fehérje glikolizáció gátlása, stb.) -vagy áthelyezni az egész rendszert egy olyan alapra, amely eleve irányítottan stacioner
Mind2 nagyon-nagyon távoli és még rengeteg mindent meg kell értenünk, de én ilyen irányokban szeretnék majd kutatni pár év múlva.
:) túl hamar álltál le vele igazából ez is csupán az ism. terj. színvonalat ütné meg valóban nem tér ki arra, h mi a GFP jelentõsége (bár sztem az ott ülõ hallgatóság nagyobb része tudja és hallott már róla épp elég elõadást, meg nem is ez volt a téma)
A vége felé még mutatott példákat az állatok és gépek "ötvözésére", távirányítású rovarok, majom, amelyik robotkart vezérel a gondolatával... no meg tápban levõ vmilyen agy, amelyiket számításokra próbálnak felhasználni
Biztos ilyen téren levõ mûveletlenségemnek köszönhetõ, h érdekes kis összefoglalónak találtam arról, h milyen dolgokat valósítottak már meg az elmúlt években. De hát vhonnan én is kell szerezzek némi ism. terj. híreket :|
Nem hiszem, h a gonosz biomérnökökrõl lett volna szó a tudósközösség egy ilyen elõadásában. Mindenesetre a végén felvetette, h etikai kérdések is felmerülnek a jövõt illetõen.
gondolom csak látványosan próbálta kihangsúlyozni hogy már ezt is lehet. Bár kitért arra hogy világító embereket is lehetne csinálni... :D csak ezt a speciális érvelést nem láttam
elõször elkezdte mutogatni a fele-fele állatokat, ekkor még reménykedtem, hogy nem arról lesz szó, hogy de gonoszak a biomérnökök/genetikusok (/stb.). aztán megemlíti a GFP-t és úgy álltja mindezt be, mint amit arra használnák, hogy olyan egereket csináljanak ami világít a sötétbe :D ez nevetséges, a molekuláris biológiai egyik legjelentõsebb módszere, amit ma már minden laborba csinálnak; a jelentõségét nem lehetne eléggé kihangsúlyozni. ekkor kapcsoltam ki a videót.
lehet, hogy nem így akarta elõadni és én értettem félre, lehet, hogy "my bad", but I felt offended.
Hallottam, h a nõk immunrendszere erõsebb/hatékonyabb, mint a férfiaké. Gondoltam utánanézek, h miért is illetve honnan ered ezen állítás. Csak ezen cikkre való hivatkozásokat találtam. Próbáltam összeolvasni, h most akkor mi is van, de el kellene a segítség az értelmezésben. Az eredeti cikkhez már nem volt türelmem, csak az elõtte levõ összefoglalókhoz/hírekhez.
Azt csipegettem össze, h az ösztrogén blokkolja a caspase-12 gént, ami gátolná, csökkentené a gyulladásos immunreakciót. Majd a caspase-12-rõl meg hogy citokinokat gyárt, amik meg pont hogy az immunreakciókat serkenti (ugye, most akkor melyik?). Aztán vmi olyanról írtak, h az afrikaiaknál van olyan változata ennek a génnek, ami túl erõs reakciókat válthatnak ki (szepszis), viszont hatékonyabbak az LPS-es (Lipopoliszacharid) mikrobák ellen.
mert én abból indultam ki, hogy az elsõ nukleotid láncba építéséhez nem kell energia (mert õ a lánc elsõ tagja), minden nukleotid a saját beépüléséhez szükséges energiáját hordozza a PPP kötésben, így a legelsõ nukleotid PPP-je megmaradna. Erre írtam, hogy a lánckezdés lehet egy sokkal bonyolultabb lépés, ami igényelheti az elsõ PPP-t, vagy alapból kilehet indulni monofoszfátból, ha nem kell + energia.
tudom, de a többi részben minden érthetõ volt:o) 1ébként a kép okés, így gondoltam én is, de itt sincsen a végtermék 1ik nukleotidján sem 3 foszfátcsoport, hanem mindhez tartozik 1 (ill, mindegyik közt van egy és egyik végén +1)
sajnos nem tudok konkrét válaszokkal szolgálni (: elképzelhetõ, hogy a méhviasz aromáját maga a viasz adja, ebben az esetben elég kicsi lehet a tenziója (egyensúlyi gõznyomása) és lehet, hogy kis koncentrációban is nagy ingert vállt ki a receptorokból, így ez megmagyarázza, hogy sokáig van intenzív illata
mert a méhviasznál gondolkodtam amikor megszagoltam, hogy év múlva is amikor odateszem magam elé intenzíven érzem az illatát, és az nem kevés anyag lehet ha folyamatosan kiereszti magából
hát ez szerintem anyagfüggõ és mérések kérdése, amire így kapásból nem tudom a választ és nem jó a sacométerem :D
pl. a kámfor nagyon gyorsan elfogy, de ez a szublimáció miatt van, ami egy fázisátalakulás. Sok aromaanyag folyadék halmazállapotú tisztán (pl citrullin,fahéjaldehid stb) ezeknek jóval nagyobb a tenziója, mint a szilárd anyagokénak de persze ez is anyagfüggõ.
Egy másik kérdésem - vannak szilárd dolgok, (pl méhviasz egy amorfabb, de van más is), ami sok év múlva is nap-nap, óra-óra után nagyon intenzív szagot ad, vagyis nyilván kiszakadnak molekulák belõle. Mennyit képes csökkenni a szilárd anyag tömege az elveszõ molekulákból?
az ok alapja az volt hogy ma már olyan megfeleltetéseken kell végigmenniük az új gyógyszereknek hogy az Aspirin nem tudna kellõen teljesíteni bizonyos pontokat
hehe, olvastam olyat is, hogy mellékhatásként kisüti az oxidatív-foszforilációt a mitokondrium membránban (ez az egyik legfontosabb energiatermelõ folyamat)
én nem hallottam arról, hogy beszeretnék tiltani (mondjuk nem is vagyok annyira up-to-date ebben a témában), de elképzelhetõ, mert az aszpirin (acetil-szalicilsav) elég aktív molekula és valószínûleg nem csak specifikusan hat (tehát sok a mellékhatása). Az én tudásom szerint (amit majd részletezni is fogok), a fájdalomcsillapításkor a prosztanoidok szintézisében résztvevõ cyclooxygenázt gátolja, méghozzá úgy, hogy az annak aktív centrumában lévõ szerin-t acilezi, ami in vivo irreverzibilis. Mivel az acetil-szalicilsav igen kicsi molekula, ezért valszeg nem annyira specifikus, sok enzim aktív centrumába befér és acilezheti az enzimet
Ha már fájdalomcsillapító: én azt hallottam elõadáson hogy ma már nem lehetne engedélyeztetni az Aspirint, ez miért van? Másrészt a kémiához kapcsolódóan remélem mindenki ismeri a Vegyészoperát, ami egyedülálló :)
Rendben, fogok írni elõször amikrõl fejbõl tudok: a cyclooxygenase inhibitorokról (pl. aszpirin) és a béta-laktám típusú antibiotikumokról (pl. penicilin-G), ha az úgy megfelel ((((:
Nagyon érdekes ez a topic. Ha megkérhetlek írnál a gyógyszerek hatásmechanizmusáról?? (pl. fájdalomcsillapítól, antibiotikumok) talán a 2. érdekelne jobban
Elóre is kössz!
viszont köszönöm a kérdést, nyugodtan írjatok bármirõl, kritikát, akármit
azt szeretném, hogy leírjam elõször az alap dolgokat, úgy, hogy az elõismeretek nélkül is érthetõ legyen (!) és csak azután írni különlegesebb dolgokról, ami a leírt alapokra támaszkodik.
pl. így hogy írtam a DNS-rõl, már írhatok a DNS-metilációról, ami engem nagyon megfogott amikor olvastam róla
értem mire gondolsz, nem írtam el (: nem tudom biztosra, hogy milyen a DNS legelsõ nukleotidja bioszintéziskor, lehetséges, hogy sokkal összetettebb mechanizmusokkal indul, én csak azt próbáltam szemléltetni, ahogy a lánchosszabbítás megy és abban trifoszfátok vannak.
Ha nincs semmilyen összetettebb lépés az elején, akkor igen, indulunk egy trifoszfáttal, jön egy második trifoszfát amirõl egy difoszfát leválik és így a dinukleotidon összesen 4 foszfátcsoport lesz: 3 a legelsõ nukleotidon és 1 a kettõ között
"Amikor két nukleozid-trifoszfát összekapcsolódik, akkor az újonnan bejövõ nukleozid-nak egy difoszfát-csoportja leválik, és a maradék foszfát-csoporton keresztül csatlakozik az elsõ nukleozid cukor részéhez(foszfodiészter kötés), így kialakítva a megszokott cukor-foszfát gerincet:" Eleinte azt mondod, h 2 nukleozid-trifoszfát kapcsolódik össze és 1 difoszfát megy el, akkor 4foszfát csop. kéne maradjon. De utána meg "az újonnan bejövõ nukleozid-nak"-ként fogalmazol, tehát akkor az eleje csak elírás volt és a helyzet az, h már van 1 foszfátcsoportos nukleotid és ahhoz kezd csatlakozni egy trifoszfátcsoportos?
1ébként nagyon rendes tõled, h ezt így összeírod, köszi.
Szeretném bemutatni az alapvetõ biomolekulákat és ezen keresztül egy alapvetõ hozzáállást szemléltetni, mely szvsz szükséges a biokémiai rendszerek (annak mechanizmusai, tulajdonságai stb.) megértéséhez. Ez egy elég nehéz feladat, mert az ember alapvetõen úgy sajátítja el, hogy huzamosabb idõn keresztül ezzel foglalkozik és kifejlõdik benne ez a speciális "intelligencia", amit elég nehéz így átadni ... mindenesetre én próbálkozom (:
A DNS (Dezoxiribonukleinsav)
Hol máshol lehetne kezdeni a biokémia tárgyalását, mint a DNS-nél, ennél a csodálatosan elegáns óriásmolekulánál, amely az élet tervrajzát szolgáltatja.[/retorikai módszerek OFF] Mindenki hallotta már, hogy a DNS-re úgy is szoktak hivatkozni, mint az örökítõanyag, genetikai információ stb., még talán az is közismert, hogy "kettõs spirál" és α-hélix. Ezeknek a fogalmaknak a megértéséhez már szükséges a molekula ismerete és a mélyebb megértéséhez (pl. hogy miért kettõs spirál) a DNS kémia tulajdonságának ismerete, amit itt be is fogok mutatni.
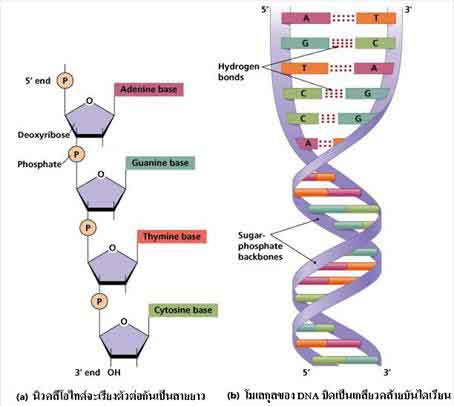
3 alapvetõ egységbõl épül fel a DNS, ahogy a képbõl is látszik: a lánc gerincét alkotó cukorból(piros 5szög) és foszfát csoportból((P)) (vagyis cukor-foszfát lánc), és a bázisokból(sárga sokszög)(amibõl 4-féle van). Ez látható az alábbi képen is a négy különbözõ bázissal:
Ha kiveszünk egyetlen bázis-cukor-foszfát 3-ast, azt nukleotidnak hívjuk és kirészletezve a kémia szerkezetét ilyen:
Ezen a képen a 4 lehetséges bázis közül az adenin van lerajzolva, ezért a fent látható molekula kémiai neve adenozin-monofoszfát. Az "-ozin" jelzi számunkra, hogy a bázishoz egy cukor is kapcsolódik, valamint a "mono-" azt jelzi, hogy 1 foszfát csoport van a molekulában, ugyanis lehetne 2 (di-), vagy 3 (tri-) is (pl. Adenozin-trifoszfát)! Ilyenkor több foszfátcsoport kapcsolódik egymás után:
Amikor a DNS bioszintézise folyik (tehát az összerakása a szervezetben), akkor az mindig 3 foszfát csoportot tartalmazó nukleotidokból történik (tehát trifoszfátok). Ez azért van, mert a nukleotidok összekapcsolása (tehát a lánc építése) energiaigényes folyamat és a foszfát-csoportok összekapcsolásában "energiát lehet tárolni". Ez úgy lehetséges, hogy egy trifoszfát molekulának a foszfát kötései viszonylag kedvezõtlenek, ezért ha létezik, akkor az magas energiájú. Így érthetõ az, hogy egy trifoszfát megbontása monofoszfáttá energiát szolgáltat a DNS bioszintéziséhez. Sõt ez annyira elterjedt, hogy a trifoszfátok (fõként az Adenozin-trifoszfát - ATP) a szervezet legfontosabb köztes-energiatároló molekulája! A legtöbb energiaigényes reakció energiáját ATP szolgáltatja (molekulák energiaigényes összerakása, molekulák energiaigényes szállítása, mozgás, beleértve az izommûködést is! stb.) és amikor a szervezetünk energiatermelõ reakciókat végez (pl glükóz lebontása), akkor az ATP formájában keletkezik, de errõl majd késõbb (:
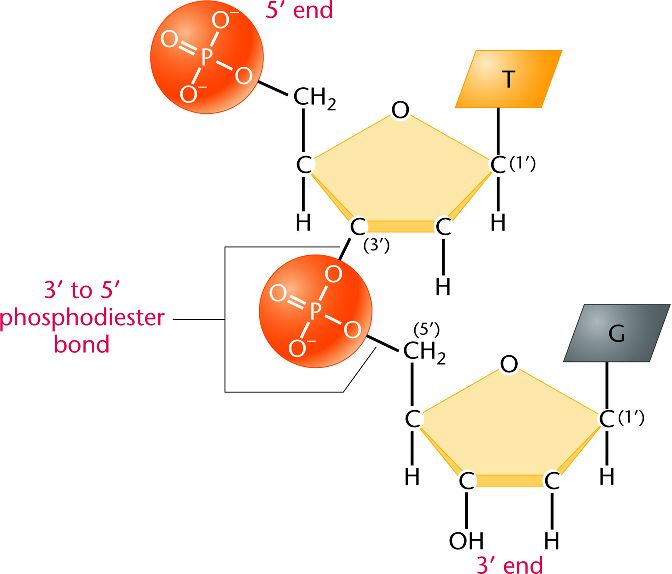
Tehát amint írtam a DNS bioszintézise nukleozid-trifosztfátokból történik, természetesen fehérjék segítségével. Amikor két nukleozid-trifoszfát összekapcsolódik, akkor az újonnan bejövõ nukleozid-nak egy difoszfát-csoportja leválik, és a maradék foszfát-csoporton keresztül csatlakozik az elsõ nukleozid cukor részéhez(foszfodiészter kötés), így kialakítva a megszokott cukor-foszfát gerincet:
A képen is fel van tüntetve a foszfátcsoport maradék negatív töltése, amely nagyon fontos szerepet tölt be a DNS szerkezetét illetõen. Nagyon fontos megérteni a hidrofobicitás/hidrofilitás fogalmát, mert nagymértékben befolyásolja a makromolekulák (pl DNS, fehérjék) térszerkezetét! Közismert, hogy vannak olyan anyagok, amelyek nem oldódnak vízben (hidrofóbok; hydros-víz, phobosz-félni), pl az olaj, míg mások igen, pl a konyhasó (nátrium-klorid, NaCl) (hidrofilek; -philia, szeretni). Az oldódás annyit jelent, hogy az oldószer molekulái (pl. víz) körbeveszik (szolvatálják, kialakul egy szolvát burok) az oldandó anyag molekuláit. Ez persze nem véletlenül történik, hanem termodinamikai hajtóereje van és csak akkor történik meg, ha a szolvatáció energianyereséggel jár. Ehhez tudnunk kell a víz szerkezetérõl, hogy egy ún. dipólusú molekula:
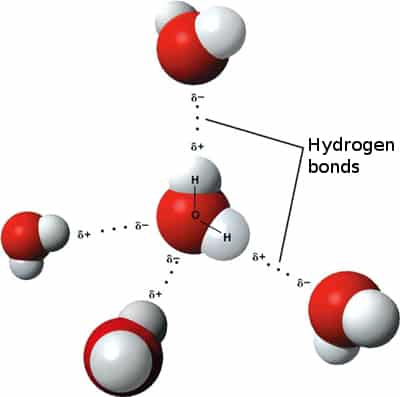
Ez annyit jelent, hogy a molekulában az elektronok nem egyenletesen oszlanak el, jelen esetben az oxigén atom felé vannak eltolva, ahogy az az elektrosztatikus potenciál képbõl is látszik (piros~nagy elektronsûrûség, kék~kicsi). Ez azért van, mert az oxigén atomnak nagyobb az elektronegativitás értéke, ami az atom szerkezetének a következménye. Képletesen szólva az oxigén magára húzza az O-H kötésben résztvevõ elektronpárt, így az oxigénen nagyobb az elektronsûrûség, ami õt picit negatívabba (δ-), míg a hidrogéneket kicsit pozitívabbá (δ+) teszi. Ennek köszönhetõen a vízmolekulák egymással ilyen ún. másodlagos kölcsönhatásokkal kapcsolódnak (dipol-dipol kölcsönhatás), ráadásul a víz molekula oxigén δ- atomja és a másik víz δ+ hidrogénje között olyan erõs kölcsönhatás lép fel, hogy az oxigén kicsit "magára húzza" a szomszédos víz hidrogénjét (hidrogén hidas kölcsönhatás)
A másodlagos kölcsönhatások csökkentik a rendszer energiáját, mert valamelyest semlegesítik egymást a kialakuló részleges töltések.
Visszakanyarodva az oldódáshoz, a NaCl egy ionos vegyület, ami azt jelenti, hogy kationokból és anionokból áll, szilárd formában még ionrácsban, amit a kilakult +/- töltések tartanak össze. Ha egy NaCl kristályt vízbe dobunk, akkor a töltéssel rendelkezõ kationokat és anionokat a dipólus H2O molekulák szolvatálni tudják, hiszen õk rendelkeznek parciális töltéssel (δ+/-), így az anyag oldódik:
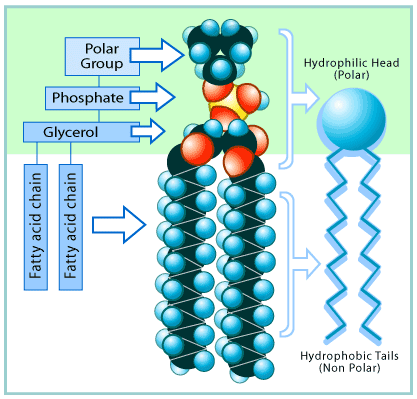
Kicsit trükkösebb a helyzet, ha egy olyan molekulánk van, aminek van hidrofób és hidrofil része is! Ilyenek a foszfolipidek, amelyek egy poláros feji részbõl (foszfát-csoport) és egy apoláros farki részbõl (hosszú szénláncú zsírsav) állnak:
Ilyenek anyagokat vízbe rakva úgy rendezõdnek molekuláris szinten, hogy a poláris hidrofil rész a vízzel, míg az apoláros hidrofób rész más apoláris részekkel érintkezzen. Ez teszi lehetõvé micellák és membránok létrejöttét:
És végül amiért ebbe az egészbe belekezdtem :D A DNS gerincét alkotó cukor-foszfát lánc poláris jellegû fõként a meglévõ negatív töltés és részlegesen a cukron lévõ, vízhez hasonló OH- csoportok miatt, míg ha a bázisokat szemrevételezzük, akkor azok fõként apolárisak, hidrofóbok. Ezért a DNS kettõs-spirál a térben úgy rendezõdik, hogy a ként láncon lévõ bázisok befele, egymással szemben -létraszerûen- helyezkednek, míg a poláris, hidrofil rész kívül van. A DNS spirális formát vesz fel, hogy minél kevésbé érintkezzenek a hidrofób bázisok vízzel.
Az így kialakult képünket a DNS-rõl még annyiban szeretném módosítani, hogy végre pontosan megnevezem a bázisokat, íme:
A,T,G,C - adenin, timin, guanin, citozin. Ez a négy bázis építi fel a DNS-t és ezeknek a sorrendje határozza meg a genetikai információt. Kettõs spirál révén, mindig két bázis van egymással szemben (bázispárosodás), mindkét szálon 1-1 nukleotidonként, így alakítva ki a létraszerû elrendezõdést. Még a hélix szerkezetének felfedezése elõtt észrevették, hogy A-ból mindig ugyanannyi van, mint T-bõl és G-bõl, mint C-bõl, ezt elnevezték Chargaff féle bázispárosodási szabálynak. Csak a szerkezet megfejtése után derült ki, hogy mindez azért van, mert párosodáskor A-val szemben mindig T van és G-vel szemben mindig C (és vica-versa). Ennek oka az, hogy a bázisok így összekapcsolódva "egymás komplementerei", a két komplementer bázis között így alakulnak ki a legerõsebb másodlagos kötések, amik stabilizálják a kettõs hélixet.
huh, ez tök jó volt, köszi! épp aktuális arra amit az elõzõ topikba írtam :D
terveim közt szerepel hamarosan írni ide a topikba az alapfogalmakról, hogy mindenki érthesse, akit érdekel a téma, és utána belemenni speciálisabb témákba (pl. a HIV-proteáz, retrovírusok)
csak nagyon kevés idõm van az egyetem miatt. talán hétvége körül írok a DNS-rõl részletesen és talán a fehérjékrõl, csak bazi nagy anyag ((:
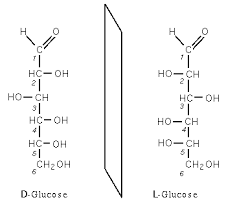
Ha van 100 db D-glükóz molekulád kristályban, akkor az nyílt láncban van, és így néz ki:
Viszont ha most ezt a 100 molekulát vízbe rakod, akkor gyûrûbe fog záródni (ez a laktonizáció), és a gyûrûnek két állása képzelhetõ el:
Ez az α,β-glükóz, pontosabban α,β glükopiranóz (ez a piranóz gyûrû). A két forma egymásba át tud alakulni oldatban, ezt hívják mutarotációnak és abból a formából lesz több, amelyik stabilabb, jelen esetben α:β 36:64 és ez azért van, mert β állásban a nagy térigényû OH- csoport a legtávolabb van a gyûrûtõl, így minimalizálódik a taszítás.
Ha létezik egy olyan molekula, amelyikben van 1 királis C-atom (tehát annak a molekulának létezik tükörképi párja, ezt úgy hívják, hogy enantiomerje) akkor az a polarizált fény síkját valamerre forgatja (+)/(-). Ha egy királis molekula a polarizált fényt (+)-ba forgatja, akkor annak a tükörképi párja (enantiomerje) (-)-ba fog forgatni. Ha összeöntöd a két molekulát, és mindkét molekulából ugyanannyi van az elegyben, akkor az már nem fog forgatni (ezt racém elegynek hívják), mert amennyit az egyik molekula (+)-ba forgat, annyit forgat a másik (-)-ba.
köszönöm szépen!:) kurva érdekes... azthiszem most egy darabig maradok is read-only módban, csak hajrá! érdekes téma, jó olvasgatni ilyeneket is
ugyanez van az aminosavakkal is, az α-C-atom királis, létezik tükörképi párja, éppen ezt raktam be az elsõ képre.
Amúgy ezt sztereokémiának hívják.
Az enzimek általában szterospecifikusak, tehát pl csak a D-glükózt tudják lebontani, ezért a természetben általában csak D cukrok és L aminosavak vannak.
igen, ez így van. Onnan érdemes megközelíteni a dolgot, hogy pl. a glükóz egy monoszacharid, ami azt jelenti, hogy 1 cukormolekula, van diszacharid(2 molek),oligoszacharid(10-es nagyságrend),poliszacharid(100-as és azon felüli nagyságrend) is. Még az is köztudott, hogy szénhidrátnak nevezzük. Ez az elnevezés onnan jön, hogy régen a molekula szerkezetét teljesen nem ismerve, azt hitték, hogy C[n](H2O)[n], tehát mintha a szénnek a hidrátja lenne valamilyen formában. Glükóz esetében n=6 és a képlete C6H12O6, vagy az elõzõre utalva C6(H2O)6. Tehát ha megnézzük akkor C,O,H-bõl áll, és amit írtál, hogy a cukrokból többféle létezik a C miatt van.
Ha van egy olyan C atom, amihez kapcsolódik 4 különbözõ másik ligandum (atom v molekula), akkor ebbõl a C atomból 2 féle létezik, melyek egymásnak tükörképei (királis), mint a két tenyér:
Ez csak akkor történik meg, ha a C-hez 4 különbözõ szubsztituens kapcsolódik, ha ezek közül 2 egyforma, akkor úgy összeforgatható a két C, hogy azok egymást fedik, tehát nem tükörképek.
Amint a képbõl is látszik, egymás tükörképe a két molekula, ugyanolyan atomok alkotják, ugyanolyan kapcsolódási sorrendben, de más térszerkezetben.Az egyiket D (dexter-jobb) a másikat L (leveus-bal) glükóznak nevezik, az alapján, hogy a királis C-atom az ún. Fischer-projekciós ábrán merre áll.
Ezek az alfa béta konformációk ha jól emlékszem, és valóban máshogy törik a fényt, és emiatt másmilyenek. Valamelyik glükózfajtából a természetben talán nincs is, de már nem emlékszek, hogy az alfa, vagy a béta változatból. Szerintem a kutatás legelejének a 10.es kémia könyv pont jó, onnan meg tudod érteni, hogy molekuláris szinten mi a különbség a két molekula között.
régebben olvastam valahol, hogy az ember (meg a szén alapú életformák a földön) csak az olyan tápanyagot (cukrot azthiszem) tud(nak) lebontani, ami bizonyos szögben töri meg a fényt... van ami jobbra, van ami balra és az egyiket nem tudja (valami alfa;béta valami volt a neve)
de nemtudom már hogy egyáltalán hogy kezdjek neki ennek az írásnak a felkutatásának, mert nagyon kulcsszót se tudok. errõl te tudsz valamit vagy vakvágányon vagyok?
elsõként legyen:
A kolera toxin patomechanizmusa
A Vibrio cholerae egy Gram-negatív baktérium, amely egy toxin hatású fehérje komplex kiválasztására képes (tehát exotoxin), ez a kolera toxin (Ctx). A Ctx felelõs a kolera tüneteiért és patomechanizmusának ismeretében választ adhatunk a tünetek kialakulásának miértjére. A patomechanizmus ismeretéhez, elõször a Ctx szerkezetét kell szemügyre venni. Az alábbi képen a Ctx sematikus szerkezete látható (PDB_ID:1xtc):
A képen látható piros (CtxA-1) és narancssárga (CtxA-2)fehérjerészlet alkotja az toxin A részét (CtxA) és a kék a B részét (CtxB). A CtxB 5 egyforma (homológ) fehérjerészbõl áll, tehát egy pentamer és egy gyûrût alkot, amelybe beleilleszkedik a CtxA-2 α-hélix része (narancssárga), a két szerkezetet egymással másodlagos kölcsönhatások révén kapcsolódnak, kovalens kölcsönhatások révén nem. A CtxA α-hélix részét (CtxA-2, narancssárga) és a késõbbiekben enzimatikus szerepû CtxA-1 részt egy cisztein-cisztein (Cys-Cys) diszulfid híd köti össze, amely a meglelõ körülmények között elszakad, szabaddá téve az enzimatikus CtxA-2 részt.
Amint az a kísérletekbõl kiderült a CtxB pentamer szerepe, hogy a sejtbe való bejutást megvalósítsa. Ez úgy történhet meg, hogy kapcsolódok a bélhámsejtek membránjában lévõ, ún. GM1 gangliozid receptorokhoz, melynek hatására a teljes fehérje komplexet bekebelezi a sejt. A kialakult membránnal körülvett struktúrát endoszómának hívjuk. Az endoszóma ezután a Golgi-készülékbe szállítódik, ahol a sejt saját fehérjéi felszakítják a CtxA-1 és CtXA-2 fehérjeláncok közötti Cys(187)-Cys(199) diszulfid kötést, ezáltal szabaddá téve az enzimatikus funkcióval bíró CtxA-1 részt.
Ezek után a CtxA-1 rész a citoplazmába jut, ahol egy másik fehérje (ún. ADP-ribolizációs faktor 6) szabaddá teszi a CtxA-1 aktív centrumát, melynek hatására az képes lesz megkötni egy NAD (Nikotinsavamind-Adenin-Dinukleotid) molekulát. Így a CtxA-1-NAD komplex kötõdik egy membránban lévõ másik fehérjéhez, egy adenilát-ciklázhoz, melnyek feladata cAMP (ciklikus-AdenozinMonoFoszfát) termelése, adott inger esetén, mely normális körülmények között a membránban lévõ G-fehérjéktõl jön. Viszont ebben az esetben a CtxA-1-NAD komplex ADP-ribozilációt hajt végre az adenilát-ciklázon, amely annyit jelent, hogy a CtXA-1-NAD komplex NAD részébõl egy ADP-ribozil részt kapcsol az adenilát-ciklázhoz, ennek eredményeként az adenilát-cikláz folyamatosan elkezd cAMP-ot termelni. A cAMP másodlagos hírvivõként szolgál általában a sejtekben, a felszaporodása egyáltalán nem rendeltetésszerû. Jelen esetben a létrejövõ hatalmas mennyiségû cAMP egy elektrolit kiáramlást fog okozni a bélhámsejtekbõl a bél lumenbe, amely víz kiáramlását vonja maga után. Ez összességében hasmenést eredményez és dehidratációhoz vezet, amely a kolera elsõ számú tünete és haláloka.
Üdvözlet minden kedves témalátogatónak ! {:
A Biokémia téma célja, hogy minden érdeklõdõ számára lehetséges legyen a biokémiáról és annak társdiszciplináiról (molekuláris biológia, sejtbiológia, bioinformatika, etc.) a fórum keretein belül megvalósuló társalgása.
Idõközönként írni fogok bõvebben az általam érdekesnek tartott témákról, amelyekrõl majd szívesen várom véleményeteket, hozzászólásaitokat és esetleges kérdéseiteket.
Bármilyen témához kapcsolódó kérdést/megnyilvánulást szívesen várunk!

 }, nem kell hozzá még ráadásként reductio ad absurdum-ja {
}, nem kell hozzá még ráadásként reductio ad absurdum-ja { }
}