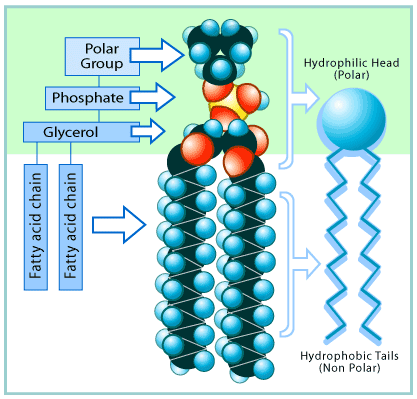
Kicsit trükkösebb a helyzet, ha egy olyan molekulánk van, aminek van hidrofób és hidrofil része is! Ilyenek a foszfolipidek, amelyek egy poláros feji részbõl (foszfát-csoport) és egy apoláros farki részbõl (hosszú szénláncú zsírsav) állnak:
Ilyenek anyagokat vízbe rakva úgy rendezõdnek molekuláris szinten, hogy a poláris hidrofil rész a vízzel, míg az apoláros hidrofób rész más apoláris részekkel érintkezzen. Ez teszi lehetõvé micellák és membránok létrejöttét:
És végül amiért ebbe az egészbe belekezdtem :D
A DNS gerincét alkotó cukor-foszfát lánc poláris jellegû fõként a meglévõ negatív töltés és részlegesen a cukron lévõ, vízhez hasonló OH- csoportok miatt, míg ha a bázisokat szemrevételezzük, akkor azok fõként apolárisak, hidrofóbok. Ezért a DNS kettõs-spirál a térben úgy rendezõdik, hogy a ként láncon lévõ bázisok befele, egymással szemben -létraszerûen- helyezkednek, míg a poláris, hidrofil rész kívül van. A DNS spirális formát vesz fel, hogy minél kevésbé érintkezzenek a hidrofób bázisok vízzel.
Az így kialakult képünket a DNS-rõl még annyiban szeretném módosítani, hogy végre pontosan megnevezem a bázisokat, íme:
A,T,G,C - adenin, timin, guanin, citozin. Ez a négy bázis építi fel a DNS-t és ezeknek a sorrendje határozza meg a genetikai információt. Kettõs spirál révén, mindig két bázis van egymással szemben (bázispárosodás), mindkét szálon 1-1 nukleotidonként, így alakítva ki a létraszerû elrendezõdést. Még a hélix szerkezetének felfedezése elõtt észrevették, hogy A-ból mindig ugyanannyi van, mint T-bõl és G-bõl, mint C-bõl, ezt elnevezték Chargaff féle bázispárosodási szabálynak. Csak a szerkezet megfejtése után derült ki, hogy mindez azért van, mert párosodáskor A-val szemben mindig T van és G-vel szemben mindig C (és vica-versa). Ennek oka az, hogy a bázisok így összekapcsolódva "egymás komplementerei", a két komplementer bázis között így alakulnak ki a legerõsebb másodlagos kötések, amik stabilizálják a kettõs hélixet.